Trying to see the invisible: what are the consequences of behavioral changes along gradients of novelty?
Abstract: Ecological novelty can be both cause and result of behavioral changes in animals. These behavioral changes can be subtle, but are likely widespread, and could have large effects on communities and ecosystems. While animal behavior can vary tremendously across space and time, I highlight three general themes regarding novelty and behavior. First, animal behavior can either create novelty, or respond to other drivers of novelty. Second, behavioral responses to novelty can either harm or benefit humans. Third, it is difficult to predict behaviors in high-novelty systems, because of the emergence of new behaviors in very novel systems. While there is considerable uncertainty surrounding the relationship of animal behavior and novelty, conducting behavioral ecology experiments across novelty gradients, and integrating known behavioral responses to novelty into forecasts of ecological change could improve predictions of the consequences of animal behavior in future habitats.
Most ecological research is based on observing and counting organisms. For example, if we know that there are 50 deer in Area A, but 10 deer in Area B, we might hypothesize that the 50 deer in Area A have a larger impact on their habitat than the 10 deer in Area B, perhaps by eating more plants or excreting more nitrogen.
However, abundance can provide a misleading measure of a population’s impact, because individuals within a species can behave very differently. In our deer example, the presence of a predator in Area A would lead deer there to spend more time watching for potential threats and less time eating. Similarly, the presence of a competitor in Area A could cause deer to avoid high-quality food patches, creating differences in diet and space use between habitats. While behavioral variation can complicate ecological monitoring and data collection, properly considering animal behavior has the potential to improve conservation efforts (Caro 2007).
Ecological novelty is not only changing where organisms are or how many organisms there are—novelty can also change how organisms behave, and behavioral novelty may precedes taxonomic novelty. Ecological novelty can alter behavior, regardless of whether populations increase, decrease, or stay the same. In turn, behavioral changes can alter community composition and ecosystem function. Below, I explore three general themes regarding ecological novelty and animal behavior, using examples from multiple study systems to illustrate the widespread links between novelty and behavior.
1: Behavior can either drive novelty or respond to novelty
Introduced animals might possess novel behaviors that rapidly change an ecosystem. For example, invasive Pacific lionfish hunt by herding their prey into corners and producing jets of water to disorient them. Prey in the native range of lionfish have evolved behavioral responses to this unique predator over many generations, but prey in the introduced range of lionfish are naïve to these tactics. Hence, prey often experience rapid population declines in habitats invaded by lionfish (Albins 2013). Alternatively, native animals may alter their behavior in response to novelty. For example, the dense growth of invasive shrubs reduces the predation risk experienced by deer, leading deer to select invaded forests over uninvaded forests. As deer aggregate under invasive shrub cover, tick survival increases, potentially posing risks to human health (Allan et al. 2010). Whether animal behavior causes novelty, or represents a response to other drivers of novelty could be an important distinction for managers trying to limit novelty. Animals whose behavior creates novelty may need to be eradicated or reduced to reach management goals, whereas animals whose behavior is a response to novelty may be left in place, if the driver of novelty is addressed.
2: Behavioral responses to novelty can either increase or decrease ecosystem services
Species vary tremendously in their functional roles and behavioral responses to novelty. In a New Mexico forest, novel noise pollution from natural gas wells reduced the efficiency of many predators, which has mixed results for human society (Francis et al. 2012). Hummingbirds respond to gas wells by increasing pollination, benefitting flowering plants. At the same time, mice respond by increasing seed predation, which limited pine regeneration. Novelty might also have no effect on animal behavior: migratory birds in Jamaica suppress insect pests even as forest conversion to agriculture intensifies (Kellermann et al. 2008). Idiosyncratic responses to novelty may frustrate land managers when they present difficult trade-offs between opposing ecosystem services. Consequently, it will be important to identify the species primarily responsible for key ecosystem services, and to quantify behavioral changes in these species along gradients of novelty. With more studies, it may be possible to generalize which services increase, decrease, or stay the same along novelty gradients.

Figure 1. Comparison of a forest understory invaded by Lonicera maackii, or bush honeysuckle (left) and an understory with the invasive shrub removed. Invasive shrubs shade out many native wildflowers and tree seedlings, but can also drastically change animal behavior, because their dense growth creates a safe refuge from predators. Photo credit: The Nature Conservancy of Kentucky
3: Behavior in high-novelty systems cannot be inferred from behavior in low-novelty systems
Some species are widely distributed along gradients of novelty, but might exhibit unique behaviors under highly novel conditions. For example, invasive Argentine ants expand their diet to include plants, creating colonies with a lower trophic position than in their native range (Tillberg et al. 2007). Similarly, an exotic predator causes waterfowl in Poland to form larger breeding colonies and nest in developed areas, behaviors that were historically rare (Brzezinski et al. 2012). These abrupt changes make it is unwise to predict behavior in highly novel habitats using data collected under low-novelty conditions.
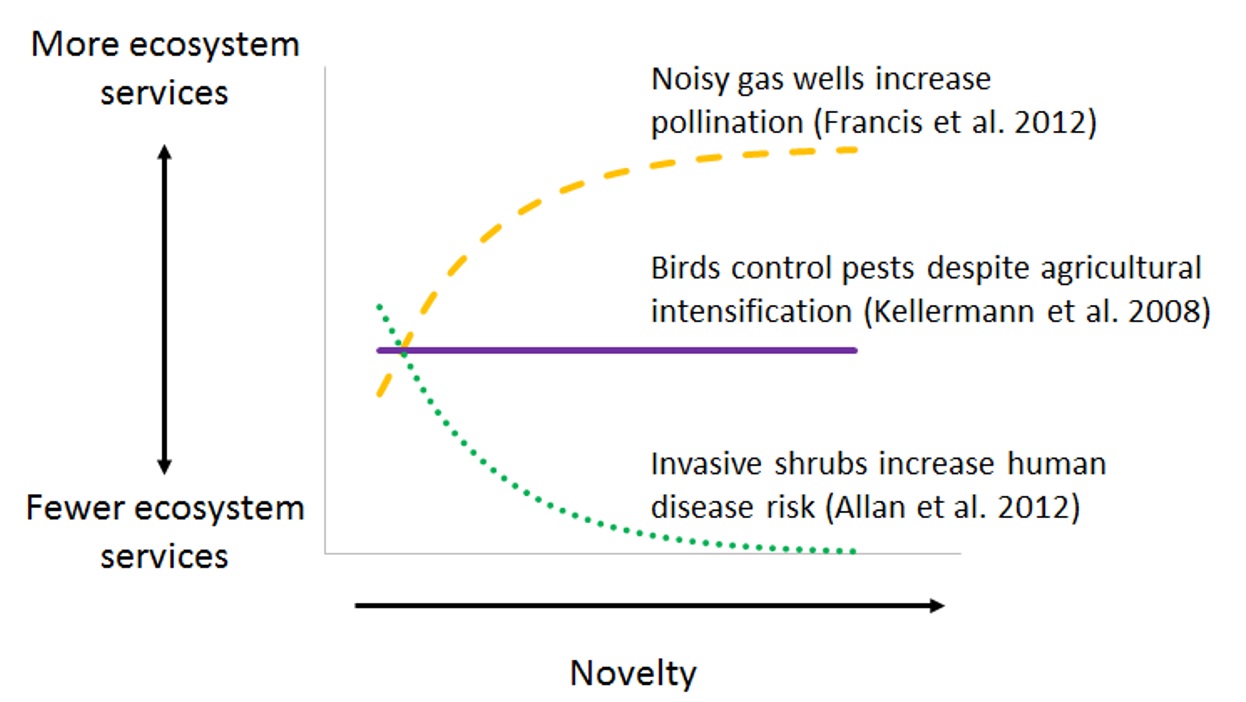
Figure 2. Animals exhibit variable responses to novelty. Because of this, as novelty increases, ecosystem services may increase, decrease, or remain constant.
Where do we go from here?
The potential for behavioral changes to produce rapid and unanticipated ecological change in highly novel systems is daunting for many ecologists and conservationists. I highlight two approaches that could be used to better understand the relationship between behavior and novelty.
1: Use experiments to measure behavioral responses to novelty
Observational data is valuable, but may not fully capture high degrees of novelty expected in future habitats. Rather, experimental manipulations may be better suited to explore behavioral responses to increased novelty, particularly when control treatments are carefully selected. Examples of possible experiments (reviewed by Knowlton and Graham 2010) include exposing organisms to novel stimuli (playback experiments), novel climates (translocation experiments), or novel habitat structure (constructing/removing corridors). Integrating behavioral experiments into research programs conducted at a large spatial or temporal extent (NEON, LTER, Nutrient Network) could be particularly valuable for advancing our understanding of how behavioral changes might influence communities and ecosystems.
- Incorporate behavioral mechanisms in species distribution models
Communities will change substantially over the next century as novelty increases. A common approach taken to predict these changes is to project species distributions onto future climate or land-use scenarios, but this approach ignores the potential for unprecedented animal behavior to emerge when novelty is high (Conley et al. 2011). Instead, a more fruitful (yet challenging) approach would be to measure behavioral changes along gradients of novelty, and then explicitly account for these behavioral changes in species distribution and disease risk models. Doing so could improve our ability to forecast the novel species assemblages of the Anthropocene, and the ecosystem services they provide.
References
Albins MA (2013) Effects of invasive Pacific red lionfish Pterois volitans versus a native predator on Bahamian coral-reef fish communities. Biol Invasions 15:29–43.
Allan BF, Dutra HP, Goessling LS, et al. (2010) Invasive honeysuckle eradication reduces tick-borne disease risk by altering host dynamics. Proc Natl Acad Sci USA 107:18523–18527.
Brzeziński M, Natorff M, Zalewski A, Z̈mihorski M (2012) Numerical and behavioral responses of waterfowl to the invasive American mink: A conservation paradox. Biol Conserv 147:68–78.
Caro T (2007) Behavior and conservation: a bridge too far? Trends Ecol Evol 22:394–400.
Conley AK, Watling JI, Orrock JL (2011) Invasive plant alters ability to predict disease vector distribution. Ecol Appl 21:329–334.
Francis CD, Kleist NJ, Ortega CP, Cruz a. (2012) Noise pollution alters ecological services: enhanced pollination and disrupted seed dispersal. Proc R Soc B Biol Sci 279:2727–2735.
Kellermann JL, Johnson MD, Stercho AM, Hackett SC (2008) Ecological and economic services provided by birds on Jamaican Blue Mountain coffee farms. Conserv Biol 22:1177–1185.
Knowlton JL, Graham CH (2010) Using behavioral landscape ecology to predict species’ responses to land-use and climate change. Biol Conserv 143:1342–1354.
McConkey KR, O’Farrill G (2015) Cryptic function loss in animal populations. Trends Ecol Evol 30:182–189.
Tillberg C V., Holway DA, Lebrun EG, Suarez A V. (2007) Trophic ecology of invasive Argentine ants in their native and introduced ranges. Proc Natl Acad Sci USA 104:20856–20861.